FREE OXYGEN AND THIYL RADICALS
Several reactive oxygen species (ROS) and one thiyl radical (RS.) are known. Among them, the most frequently studied are given below.
This ROS is formed when oxygen takes up one electron and as leaks in the mitochondrial electron transport but its formation is easily increased when exogenous components (redox cycling compounds) are present. Its first production site is the internal mitochondrial membrane (NADH ubiquinone reductase and ubiquinone cytochrome C reductase). This species is reduced and forms hydrogen peroxide (H2O2). The production of superoxide radicals at the membrane level (NADPH oxidase) is initiated in specialized cells (oxidative burst) with phagocytic functions (macrophages) and contributes to their bactericid action. The flavin cytosolic enzyme xanthine oxidase found in quite all tissues and in milkfat globules generates superoxide radicals from hypoxanthine and oxygen and is supposed to be at the origin of vascular pathologies.
While superoxide radical can be directly toxic (Fridovich I, Arch Biochem Biophys 1986, 247, 1), it has a limited reactivity with lipids, raising questions about its true toxicity. Thus, its action is frequently considered to result from a secondary production of the far-more reactive .OH species by the iron-catalyzed Haber-Weiss reaction. As that production may be limited in vivo, it was proposed that nitric oxide reacting with O2.- generates secondary cytotoxic species (peroxinitrite anion).
Hydrogen peroxide is mainly produced by enzymatic reactions. These enzymes are located in microsomes, peroxysomes and mitochondria. Even in normoxia conditions, the hydrogen peroxide production in relatively important and leads to a constant cellular concentration between 10-9 and 10-7 M.
In plant and animal cells, superoxide dismutase is able to produce H2O2 by dismutation of O2.- , thus contributing to the lowering of oxidative reactions. The natural combination of dismutase and catalase contributes to remove H2O2 and thus has a true cellular antioxidant activity. H2O2 is also able to diffuse easily through cellular membranes.
In the presence of Fe2+, H2O2 produces the very active species .OH by the Fenton reaction (described in 1894):
Fe2+ + H2O2 —-> Fe3+ + .OH + OH–
This iron-catalyzed decomposition of oxygen peroxide is considered the most prevalent reaction in biological systems and the source of various deleterious lipid peroxidation products.
It must be noticed that an important part of hydroxyl radicals is also produced (with .NO2) by the decay of peroxinitrite or peroxynitrous acid
Another reaction involving myeloperoxidase and Cl– ions represent an important .OH production process in neutrophils during phagocytosis.
Nitric oxide is produced in various types of cells and is well studied in vascular endothelium. While this species is not too reactive (poorly oxidizing function), even antioxidant under physiological concentrations (up to 100 nM), it reacts rapidly with oxygen to yield nitrogen dioxide (.NO2) which in turn may react with .NO to yield nitrogen trioxide (N2O3). The rapid reaction of O2.-, produced in different pathological states, with .NO gives the extremely reactive peroxinitrite (ONOO–) which mediates oxidation, nitrosation, and nitration reactions. In alkaline solutions, ONOO– is stable but decays rapidly once protonated into peroxynitrous acid. Its decomposition forms .OH and nitrogen dioxide (NO2.), radicals important in the formation of acid rain. The high rate and broad distribution of .NO production, combined with its facile reactions with oxygen radical species, assure that it plays a central role in regulating oxidant reactions.
Multiple mechanisms account for the nitration of lipids by .NO-derived species (O’Donnell V B et al., Circ Res 2001, 88, 12). In acidic conditions, protonation of NO2– to HNO2 can mediate the nitration of polyunsaturated fatty acids and lipid hydroperoxides giving nitrated lipid products whose structure and function are incompletely characterized.
Nitric oxide is naturally formed in activated macrophages and endothelial cells and is considered as an active agent in several pathologies based on inflammation, organ reperfusion and also may play an important role in atherosclerosis.
This chemical form of oxygen is not a true radical but is reported to be an important ROS in reactions related to ultraviolet exposition (UVA, 320-400 nm). Its toxicity is reinforced when appropriate photoexcitable compounds (sensitizers) are present with molecular oxygen. Several natural sensitizers are known to catalyzed oxidative reactions such as tetrapyrroles (bilirubin), flavins, chlorophyll, hemoproteins and reduced pyridine nucleotides (NADH). Some of these sensitizers are also found in foods and cosmetics. Some others are used for therapeutic purposes (anticancer treatments) and are sensitive to visible light. The presence of metals contributes to increase the production of singlet oxygen, as well as anion superoxide, and thus accelerates the oxidation of unsaturated lipids generating hydroperoxides.
It has been suggested that singlet O2 may be formed during the degradation of lipid peroxides and thus may cause the production of other peroxide molecules. This singlet O2 formation may account for the chemiluminescence observed during lipid peroxidation.
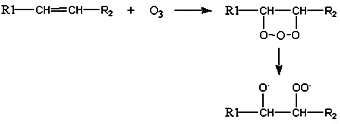
This natural compound present in the higher atmosphere and in the lower atmosphere of our polluted cities is a major pollutant formed by photochemical reactions between hydrocarbons and nitrogen oxides. Ozone is not a free radical but, as singlet oxygen, may produce them, stimulates lipid peroxidation and thus induces damages at the lipid and protein levels in vivo mainly in airways.
The exact chemistry of ozone-mediated stimulation of peroxidation is not entirely known. Ozone may add on across a double bond and decomposes to form a free radical. The proposed mechanism is given below.
Aliphatic thiols (RSH) are contained in living organisms in high concentrations. Typical levels of intracellular of glutathione are about of 5 to 10 mM. Furthermore, the level of protein SH may exceed that of GSH. The thiolate specie RS– is one of the most reactive functional groups found in proteins. It can react as a nucleophile and attack a disulfide bond. In the absence of oxygen, a thiyl radical was shown to induced cis/trans-isomerization of linoleic acid and led to several isomers. (Schwinn J, Int J Radiat Biol 1998, 74, 359). A review of the effects of thiyl radicals on lipid structures and metabolisms may be consulted (Ferreri C et al., Cell Mol Life Sci 2005, 62, 834).
Thiol compounds (RSH) are frequently oxidized in the presence of iron or copper ions:
RSH + Cu2+ —-> RS. + Cu+ + H+
These thiyl radicals have strong reactivity in combining with O2 :
RS. + O2 —> RSO2.
Furthermore, they are able to oxidize NADH into NAD. , ascorbic acid and to generate various free radicals (.OH and O2.-). These thiyl radicals may also be formed by homolytic fission of disulfide bonds in proteins.
The formation of these reactive free radical is observed in cells treated with CCl4 . The action of the cytochrome P450 system generates the trichloromethyl radical (.CCl3) which is able to react with oxygen to give several peroxyl radicals (i.e. .O2CCl3).
An intrinsic problem in the determination of rate constants in lipid oxidation is the uncertainty about the rate of initiation Ri, according to the reaction:
RH + X• ⇒ R• + XH
One possible way of overcoming this problem is to introduce into the reaction mixture a compound that decomposes at a constant rate to free radicals (X•) capable of extracting a hydrogen atom from the unsaturated lipids and consequently initiating the autoxidation process. The compounds most frequently used for this are the so-called azo-initiators (X–N=N=X), which thermally decompose to highly reactive carbon-centered radicals (Liegeois C et al., 2000, J Agric Food Chem 48, 1129). Therefore, azo-initiators are useful for in vitro studies of lipid peroxidation generating free
radicals according to the following reaction:
X−N=N−X ⇒ 2X• + N2
The water-soluble azo-initiator AAPH[2,2Azo-bis(2-amidinopropane)dihydrochloride] can be used to produce radicals in the aqueous phase, whereas the lipid-soluble AMVN [2,2′azo-bis-(2,4-dimethylvaleronitrile)] can be used to produce radicals in the lipid phase. AAPH decomposes with a first-order rate constant of Kd = 6.6 × 10−5/min at 37°C, and the flux of the free radicals is proportional to the AAPH concentration (Niki E, Chem Phys Lipids 1987, 44, 227).
DISPERSIVE LIQUID-LIQUID MICROEXTRACTION
Lire la suiteDevenez membre et participez au développement de la Lipidomique au XXIème siècle.
S'inscrire