
DICARBOXYLIC ACIDS
Although the dicarboxylic acids do not occur in appreciable amounts as components of animal or vegetal lipids, they are in general important metabolic products of fatty acids since they originate from them by oxidation. Dicarboxylic acids are suitable substrates for preparation of organic acids for the pharmaceutical and food industries. Furthermore, they are useful materials for the preparation of fragrances, polyamides, adhesives, lubricants, and polyesters.
They have the general type formula
HOOC-(CH2)n-COOH
In vegetal, a great variety of molecular forms of dicarboxylic acids are found :
simple forms with a straight carbon chain or a branched chain
complex forms with a dicarboxylic acid and an alkyl side chain : alkylitaconates
1 – Simple forms of dicarboxylic acids
Short-chain dicarboxylic acids are of great importance in the general metabolism and up to n=3 they cannot be considered as lipids since their water solubility is important. The simplest of these intermediates is oxalic acid (n=0), the others are malonic (n=1), succinic (n=2) and glutaric (n=3) acids.
The other lipid members of the group found in natural products or from synthesis have a "n" value from 4 up to 21.
Adipic acid (n=4) : Despite its name (in Latin adipis is fat), this acid (hexanedioic acid) is not a normal constituent of natural lipids but is a product of oxidative rancidity (lipid peroxidation). It was obtained by Dieterle W et al. (Ber. 1884, 17, 2221) by oxidation of castor oil with nitric acid (splitting of the carbon chain close to the OH group). Synthesized in 1902 from tetramethylene bromide, it is now obtained by oxidation of cyclohexanol or cyclohexane. It has several industrial uses in the production of adhesives, plasticizers, gelatinizing agents, hydraulic fluids, lubricants, emollients, as an additive in the manufacture of some form of nylon (nylon-6,6), polyurethane foams, leather tanning, urethane and also as an acidulant in foods. Adipic acid is used after esterification with various groups such as dicapryl, di(ethylhexyl), diisobutyl, and diisodecyl.
A graphic chart describing the biosynthesis of adipic acid via omega oxidation may be found on the BioCarta web site.
Pimelic acid (n=5) : this acid (heptanedioic acid), from the Greek pimelh (pimele fat), as adipic acid, was isolated from oxidized fats. It was obtained in 1884 by Ganttner F et al. (Ber. 1884, 17, 2212) as a product of ricinoleic acid (hydroxylated oleic acid) from castor oil.
Suberic acid (n=6) : it was firstly produced by nitric acid oxidation of cork (Latin suber) material and then from castor oil (Tilley TG, Ann 1841, 39, 160). The oxidation of ricinoleic acid produces, by splitting at the level of the double bond and at the level of the OH group, at the same time, suberic acid (octanedioic acid) and the next homologue azelaic acid.
Suberic acid was used in the manufacture of alkyd resins and in the synthesis of polyamides leading to nylon.
Azelaic acid (n=7) : nonanedioic acid is the best known dicarboxylic acid. Its name stems from the action of nitric acid (azote, nitrogen, or azotic, nitric) oxidation of oleic or elaidic acid. It was detected among products of rancid fats (Nicolet BH et al., J Ind Eng Chem 1916, 8, 416 and Nunn L et al., Biochem J 1938, 32, 1974). Its origin explains for its presence in poorly preserved samples of linseed oil and in specimens of ointment removed from Egyptian tombs 5000 years old (Banks A et al., Analyst 1933, 58, 265). Azelaic acid was prepared by oxidation of oleic acid with potassium permanganate (Ganttner F et al., Ber. 1881, 14, 1545), but now by oxidative cleavage of oleic acid with chromic acid or by ozonolysis.
Azelaic acid is used, as simple esters or branched-chain esters) in the manufacture of plasticizers (for vinyl chloride resins, rubber), lubricants and greases. Azelaic acid is now used in cosmetics (treatment of acne). It displays bacteriostatic and bactericidal properties against a variety of aerobic and anaerobic micro-organisms present on acne-bearing skin. Azelaic acid was identified as a molecule that accumulated at elevated levels in some parts of plants and was shown to be able to enhance the resistance of plants to infections (Jung HW et al., Science 2009, 324, 89).
Sebacic acid (n=8) : decanedioic acid was named by Thenard LJ (1802) from the Latin sebaceus (tallow candle) or sebum (tallow) in reference to its use in the manufacture of candles. Thenard LJ isolated this compound from distillation products of beef tallow. In 1954, it was reported that it was produced in excess of 10000 tons annually by alkali fission of castor oil (Kadesch RG, J Am Oil Chem Soc 1954, 31, 568).
Sebacic acid and its derivatives, as azelaic acid, have a variety of industrial uses as plasticizers, lubricants, diffusion pump oils, cosmetics, candles, etc. It is also used in the synthesis of polyamide, as nylon, and of alkyd resins.
An isomer, isosebacic acid, has several applications in the manufacture of vinyl resin plasticizers, extrusion plastics, adhesives, ester lubricants, polyesters, polyurethane resins and synthetic rubber.
Dodecanedioic acid (n=10) : that acid is used in the production of nylon (nylon-6,12), polyamides, coatings, adhesives, greases, polyesters, dyestuffs, detergents, flame retardants, and fragrances. It is now produced by fermentation of long-chain alkanes with a specific strain of Candida tropicalis (Kroha K, Inform 2004, 15, 568). Its monounsaturated analogue (traumatic acid) is described below.
Suberic acid, nonanedioic acid (and its mono-unsaturated derivative: 2-nonenedioic acid), decanoic acid (and its mono-unsaturated derivative: 2-decenedioic acid) are present in honey (Schievano et al., J Agric Food Chem 2013, 61, 1747) and are of special interest because they have long been recognized as part of the pheromone system of the honeybee Apis mellifera (Lercker G et al., Lipids 1981, 16, 912). They are produced in the mandibular glands of the queen and the worker honeybees, they regulate their activities in the hive.
It was shown that all these dicarboxylic acids are formed during the drying process of paint oils and that the determination of these decomposition products may be of value in determining the age of old samples.
The higher weight dicarboxylic acids (n=10 to 21) are found in different plant lipids, particularly in what was named erroneously Japan wax (triglycerides containing C20, 21, 22 and 23 dicarboxylic acids besides normal fatty acids) from the sumach tree (Rhus sp.). Among them, Thapsic acid (n=14) was isolated from the dried roots of the Mediterranean "deadly carrot", Thapsia garganica (Umbelliferae), but others, as Brassylic acid (n=11), were prepared chemically from different sources.
Brassylic acid can be produced chemically from erucic acid by ozonolysis but also by microorganisms (Candida sp) from tridecane. This diacid is produced on a small commercial scale in Japan for the manufacture of fragrances.
A review on the applications and the industrial biotechnology of these molecules has been released by Kroha K (Inform 2004, 15, 568).
A large survey of the dicarboxylic acids present in Mediterranean nuts revealed unusual components (Dembitsky VM et al., Food Chem 2002, 76, 469). A total of 26 minor acids (from 2 in pecan to 8% in peanut) were determined : 8 species derived from butanedioic acid, likely in relation with photosynthesis, and 18 species with a chain from 5 to 22 carbon atoms.
Higher weight acids (>C20) are found in suberin present at vegetal surfaces (outer bark, root epidermis). C16 to C26 a,w-dioic acids are considered as diagnostic for suberin. With C18:1 and C18:2, their content amount from 24 to 45% of whole suberin. They are present at low levels (< 5%) in plant cutin, except in Arabidopsis where their content can be higher than 50% (Pollard Met al., Tr Plant Sci 2008, 13, 236).
The first allenic dicarboxylic acid, named glutinic acid (2,3-pentadienedioic acid) was isolated from Alnus glutinosa (Betulaceae) (Hans EA, Berich Deut Chem Ges 1908, 40, 4760).
It was shown that hyperthermophilic microorganisms specifically contained a large variety of dicarboxylic acids (Carballeira NM et al., J Bacteriol 1997, 179, 2766). This is probably the most important difference between these microorganisms and other marine bacteria. Dioic fatty acids from C16 to C22 were found in an hyperthermophilic archaeon, Pyrococcus furiosus. Short and medium chain (up to 11 carbon atoms) dioic acids have been discovered in Cyanobacteria of the genus Aphanizomenon (Dembitsky VM et al., Biochemistry (Moscow) 2001, 66, 72).
A monounsaturated dicarboxylic acid, traumatic acid, (10E-dodeca-1,12-dicarboxylic acid), was among the first biologically active molecules isolated from plant tissues (English J et al., Science 1939, 90, 329). That dicarboxylic acid was shown to be a potent wound healing agent in plant that stimulates cell division near a wound site (Farmer EE, Plant Mol Biol 1994, 26, 1423), it derives from 18:2 or 18:3 fatty acid hydroperoxides after conversion into oxo fatty acids.
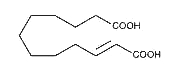
Traumatic acid
While polyunsaturated fatty acids are unusual in plant cuticles, a diunsaturated dicarboxylic acid has been reported as a component of the surface waxes or polyesters of some plant species. Thus, octadeca-c6,c9-diene-1,18-dioate, a derivative of linoleic acid, is present in Arabidopsis and Brassica napus cuticle (Bonaventure G et al., Plant J 2004, 40, 920).
Dicarboxylic acids were shown in 1934 to be produced by
w-oxidation of fatty acids during their catabolism (Verkade PE et al., Biochem J 1934, 28, 31). Thus, these authors have discovered that these compounds appeared in urine after administration of tricaprin and triundecylin. Although the significance of their biosynthesis remains poorly understood, it was demonstrated that w-oxidation occurs in rat liver but at a low rate, needs oxygen, NADPH and cytochrome P450. It was later shown that this reaction is more important in starving or diabetic animals where 15% of palmitic acid is subjected to w-oxidation and then to b-oxidation (Wada F et al. Biochim Biophys Acta 1977, 487, 261), this generates malonyl-coA which is further used in saturated fatty acid synthesis.
It was proposed recently that dicarboxylic acids are alternate lipid substrates in parenteral nutrition (Greco AV et al., Clin Nutr 1995, 14, 143). Basically, they are water soluble, undergo b-oxidation, do not induce ketogenesis but rather promote gluconeogenesis. They could represent an immediately available form of energy. Thus, inorganic salts of sebacic (C10) and dodecanedioic (C12) acids were firstly proposed, but now, triglycerides containing these fatty acids are under investigation (Capristo E et al. Clin Chim Acta 1999, 289, 11). Treatment of rats with derivatives of C16 dioic acid have shown that this compound markedly improved lipid metabolism (Russell JC et al., Arterioscl Thromb 1991, 11, 602) and inhibited the development of advanced cardiovascular disease (Russell JC et al., Arteriocl Thromb Vasc Biol 1995, 15, 918).
It must be recalled that the determination of the dicarboxylic acids generated by permanganate-periodate oxidation of monoenoic fatty acids was useful to study the position of the double bond in the carbon chain (Longmuir KJ et al., Anal Biochem 1987, 167, 213).
Long-chain dicarboxylic acids containing vicinal dimethyl branching near the centre of the carbon chain have been discovered in the genus Butyrivibrio, bacteria which participate in the digestion of cellulose in the rumen (Klein RA et al., Biochem J 1979, 183, 691). These fatty acids, named diabolic acids, have a chain length depending on the fatty acid used in the culture medium. The most abundant diabolic acid in Butyrivibrio had a 32-carbon chain length.
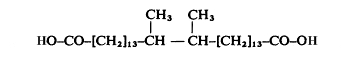
Diabolic acid (15,16-dimethyltriacontanedioic acid)
These diacids were also detected in he core lipids of the genus Thermotoga of the order Thermotogales, bacteria living in solfatara springs, deep-sea marine hydrothermal systems and high-temperature marine and continental oil fields (Huber R et al., Arch Microbiol 1986, 144, 324). It was shown that about 10% of their lipid fraction were symmetrical C30 to C34 diabolic acids. The C30 (13,14-dimethyloctacosanedioic acid) and C32 (15,16-dimethyltriacontanedioic acid) diabolic acids have been described in Thermotoga maritima (Caballeira NM et al., J Bacteriol 1997, 179, 2766).
Some parent C29 to C32 diacids but with methyl groups on the carbons C-13 and C-16 have been isolated and characterized from the lipids of thermophilic anaerobic eubacterium Themanaerobacter ethanolicus (Jung S et al., J Lipid Res 1994, 35, 1057). The most abundant diacid was the C30 a,w-13,16-dimethyloctacosanedioic acid.
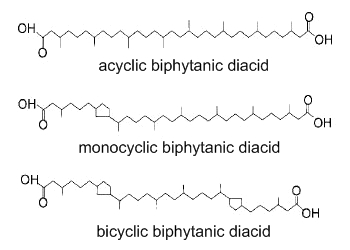
Biphytanic diacids are present in geological sediments and are considered as tracers of past anaerobic oxidation of methane (Birgel D et al., Org Geochem 2008, 39, 152). Several forms without or with one or two pentacyclic rings have been detected in Cenozoic seep limestones. These lipids may be unrecognized metabolites from Archaea.
Crocetin is the core compound of crocins (crocetin glycosides) which are the main red pigments of the stigmas of saffron (Crocus sativus) and the fruits of gardenia (Gardenia jasminoides). Crocetin is a 20-carbon chain dicarboxylic acid which is a diterpenenoid and can be considered as a carotenoid. It was the first plant carotenoid to be recognized as early as 1818 while the history of saffron cultivation reaches back more than 3,000 years.
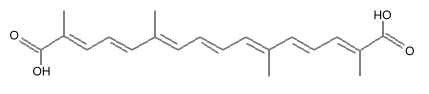
Crocetin
The major active ingredient of saffron is the yellow pigment crocin 2 (three other derivatives with different glycosylations are known) containing a gentiobiose (disaccharide) group at each end of the molecule.
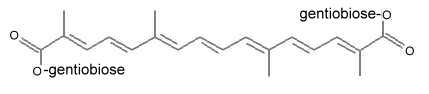
Crocin
A simple and speccific HPLC-UV method has been developed to quantify the five major biologically active ingredients of saffron, namely the four crocins and crocetin (Li N et al., J Chromatogr A 1999, 849, 349).
2 – Alkylitaconates
Several dicarboxylic acids having an alkyl side chain and an itaconate core have been isolated from lichens and fungi, itaconic acid (methylenesuccinic acid) being a metabolite produced by filamentous fungi.
Among these compounds, several analogues, called chaetomellic acids with different chain lengths and degrees of unsaturation have been isolated from various species of the lichen Chaetomella (two of them are shown below).
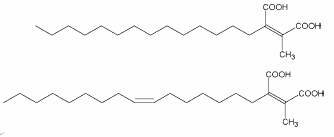
These molecules were shown to be valuable as basis for the development of anticancer drugs due to their strong farnesyltransferase inhibitory effects (Singh SB et al., Bioorg Med Chem 2000, 8, 571).
In 1999, a series of new fungal alkyl- and alkenyl-itaconates, ceriporic acids, were found in cultures of a selective lignin-degrading fungus (white rot fungus), Ceriporiopsis subvermispora (Enoki M et al., Chem Lett 2000, 54-55, Amirta R et al., Chem Phys Lipids 2003, 126, 121). Two of them are shown below.
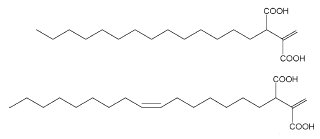
It was determined that these ceriporic acids suppressed iron redox reactions to attenuate OH production by the Fenton reaction in the presence of iron reductants such as hydroquinone and cysteine (Enoki M et al., Chem Phys Lipids 2002, 120, 9). It was proposed that the suppression of the cellulolytic active oxygen species, OH, by this metabolite contributes to the selective lignin-degradation with a minimum loss of cellulose. The absolute configuration of ceriporic acids, their stereoselective biosynthetic pathway and the diversity of their metabolites have been largely discussed (Nishimura H et al., Chem Phys Lipids 2009, 159, 77).
FATTY ACID CARBONATES
Carbonates (esters of carbonic acid, H2CO3) are well known to chemists as they represent an important class of organic compounds and among them oleochemical carbonates have interesting characteristics which make them candidates for many industrial applications.
The most common carbonates have the following structure : RO-CO-OR
R is a linear chain with 8 to 18 carbon atoms, saturated or with one double bond (dioleyl carbonate), or a branched chain (ethylhexyl, butyloctyl or hexyldecyl).
They are miscible in organic solvents but insoluble in water. Unsaturation or branching on the alkyl chain lowers their melting point (Kenar JA, Inform 2004, 15, 580).
The condensation of phosgene (ClCOCl) with an alcohol appears the most commonly used procedure to synthesize oleochemical carbonates.
The polar nature of the carbonate moiety enables it to adhere strongly to metal surfaces. Thus, they are used as lubricant components which have a protective property for metal corrosion. Some C8 to C18 carbonates have been exploited in personal-care products (sunscreen, cosmetics), dioctyl carbonate being also used as emollient or solvent in UV-filter solutions.
Extraction of metal ions (gold, silver, platinum) is improved by the use of the chelating properties of oleochemical carbonates when mixed with the metal-containing aqueous phase. Future developments will ensure a growing interest in these molecules.
PHENYL AND BENZOIC ALKANOIC ACIDS
Short chain w-phenylalkanoic acids have long been known to occur in natural products.
The simplest one is benzoic acid. This acid is present, free or esterified, in several plants.
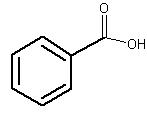
Benzoic acid
Appreciable amounts have been found in most berries (around 0.03 to 0.15%). Ripe fruits of several Vaccinium species (Ericaceae) (e.g., cranberry, Vaccinium vitis idaea; bilberry, Vaccinium macrocarpon) contain the highest concentration of free benzoic acid. Gum benzoin, a balsamic resin produced by shrubs (Styrax species) from tropical regions, mainly in Asia, contains up to 20% of benzoic acid and 40% benzoic acid esters. The name "benzoic" is derived from the name of these resins. In perfumery, benzoin is used as a fixative, slowing the dispersion of essential oils into the air.
Phenylacetic, 3-phenylpropanoic and 3-phenylpropenoic (cinnamic) acids are present in propolis, mammalian exocrine secretions or plant fragrances.
Cinnamic acid is obtained from cinnamon or from balsam. It is also found in shea butter. It is used as a component of several flavors and certain pharmaceuticals (as methyl, ethyl or benzyl esters).
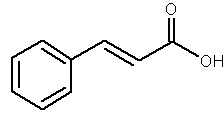
Cinnamic acid
Cinnamic acid is formed biosynthetically from phenylalanine and is metabolized through hydroxylation and methylation into various polyphenols with antioxidant properties (caffeic, coumaric, ferulic and sinapic acids). Considering their high polarity, these compounds cannot be considered as true lipids.
During a systematic study of the lipids from seeds of the plant Araceae, Schmid PC et al. (Phytochemistry 1997, 45, 1173) discovered the presence of 13-phenyltridecanoic acid as a major component (5-16% of total fatty acids). Other similar compounds but with 11 and 15 carbon chain lengths and saturated or unsaturated were shown to be also present but in lower amounts. At the same time, the even carbon chain w-phenylalkanoic acids of C10 up to C16 were discovered in halophilic bacteria (Caballeira NM et al., Lipids 1997, 32, 1271).
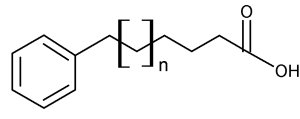
w-phenylalkanoic acid (x = 1 to 17)
Later, an exhaustive study of 17 genus of the subfamily Aroideae of Araceae revealed the presence of three major acids, 11-phenylundecanoic acid, 13-phenyltridecanoic acid and 15-phenylpentadecanoic acid in seed lipids (Meija J et al., Phytochemistry 2004, 65, 2229). Other odd carbon number acids from C7 to C23 were detected but in trace amounts. Similarly, two series of homologous odd carbon number monounsaturated w-phenylalkanoic acids were found.
Thus, it can be stated that all odd carbon chain w-phenylalkanoic acids from C1 through C23 have been found in nature. Furthermore, even carbon chain w-phenylalkanoic acids from C10 through C16 were also detected.
Two peroxidized isomeric phenylhexadecanoic acids, epiplakinic acids, have been isolated from the Palauan sponge Plakortis nigra (Sandler JS et al., J Nat Prod 2002, 65, 1258). One of them is shown below. Both acids inhibited the HCT-116 human colon tumor cell line.
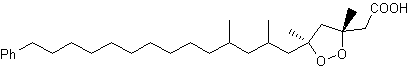
Epiplakinic acid
Substituted phenylalkenoic acids are periodically encountered in nature. As an example, rubrenoic acids were purified from Alteromonas rubra, compounds which showed bronchodilatatoric properties (Holland GS et al., Chem Ind 1984, 850).
Methyl phenylalkenoic acids (5 carbon chain) have been described from a terrestrial Streptomycete (Mukku VJ et al., Z Naturforsch 2002, 57b, 335).
Serpentene, a similar polyunsaturated phenylalkenoic acid, is also produced by Streptomyces and was shown to have some antibacterial properties.
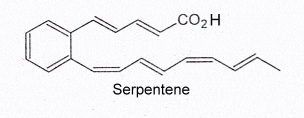
Several serpentene-like compounds have also been isolated from the same bacterial source (Wenzel SC et al., J Nat Prod 2004, 67, 1631).
Several bicyclic derivatives of linolenic acid were shown to be generated by alkali isomerization (Matikainen J et al., Tetrahedron Lett 2003, 59, 567).
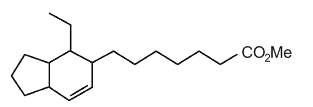
Bicyclic hexahydroindenoic acid
Some others (alkyl-phenyl)-alkanoic acids) are formed when linolenic acid is warmed at 260270°C (Hase A et al., JAOCS 1978, 55, 407).
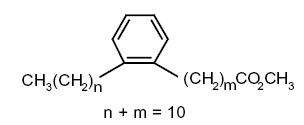
Several forms with 20 or 22 carbon atoms were identified in archaeological pottery vessels and were presumed to have been generated during heating of highly unsaturated fatty acids. They were used as biomarkers to trace the ancient processing of marine animal in these vessels (Craig OE et al., PNAS 2011, 108, 17910).
Several benzoic acid derivatives have been described in leaves of various Piperaceae species. Thus, a prenylated benzoic acid acid derivative, crassinervic acid, has been isolated from Piper crassinervium (Lago JH et al., J Nat Prod 2004, 67, 1783).
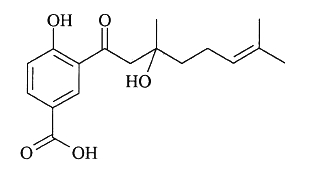
Crassinervic acid
Similar compounds were isolated from P. aduncum (aduncumene) and P. gaudichaudianum (gaudichaudianic acid). All these molecules showed high potential as antifungal compounds. A prenylated benzoic acid with a side chain formed of two isoprene units has also been isolated from the leaves of P. aduncum (Baldoqui DC et al., Phytochemistry 1999, 51, 899). More recently, three prenylated benzoic acid derivatives with four isoprene units have been extracted from the leaves of Piper heterophyllum and P. aduncum (Flores N et al., Phytochemistry 2009, 70, 621). These compounds displayed moderate antiplasmodial (against Plasmodium falciparum) and trypanocidal (against Trypanosoma cruzi) activities.
FATTY ACYL-CoA ESTERS
These fatty acid derivatives may be considered as complex lipids since they are formed of one fatty acid, a 3′-phospho-AMP linked to phosphorylated pantothenic acid (vitamin F) and cysteamine. However, to simplify the nomenclature and taking into account their metabolism, we classify them within the big group of the fatty acids and their simple derivatives rather than within the complex and phosphorylated lipids.
Long-chain acyl-CoA esters are substrates for a number of important enzymatic reactions and play a central role in the regulation of metabolism as allosteric regulators of several enzymes. To participate in specific metabolic processes, fatty acids must first be activated by being joined in thioester linkage (R-CO-SCoA) to the -SH group of coenzyme A. The thioester bond is a high energy bond.
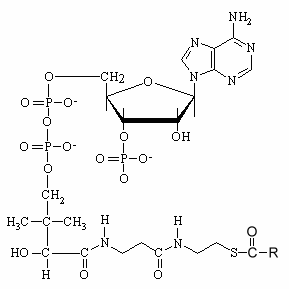
R = fatty carbon chain
The activation reaction normally occurs in the endoplasmic reticulum or the outer mitochondrial membrane. This is an ATP-requiring reaction (fatty acyl-CoA synthase), yielding AMP and pyrophosphate (PPi). Different enzymes are specific for fatty acids of different chain length.
Then, the acyl CoA esters are transported in mitochondria. They are converted to fatty acyl carnitine by carnitine acyl transferase I, an enzyme of the inner leaflet of the outer mitochondrial membrane. Fatty acyl carnitine is then transported by an antiport in exchange for free carnitine to the inner surface of the inner mitochondrial membrane. There carnitine acyl transferase II reverses the process, producing fatty acyl-CoA and carnitine. This shuttle mechanism is required only for longer chain fatty acids.
Once inside the mitochondrial matrix, the fatty acyl–CoA derivatives are degraded by a series of reactions that release acetyl-CoA and leads to the production of NADH and FADH2. There are four steps in fatty acid oxidation pathway; oxidation, hydration, oxidation, and thiolysis. It requires 7 rounds of this pathway to degrade palmitate (a C16 fatty acid).
A graphic chart of these important metabolic steps may be found in the BioCarta web site.
DIVINYL ETHER FATTY ACIDS
Fatty acid hydroperoxides generated by plant lipoxygenases from linoleic and linolenic acids are known to serve as substrates for a divinyl ether synthase which produces divinyl ether fatty acids. Up to date divinyl ethers were detected only within the plant kingdom.
The discovery of that class of compounds dates back to 1972, when Galliard T et al. described the structures of two ether C18 fatty acids generated by homogenates of the potato tuber (Galliard T et al., Biochem J 1972, 129, 743). These compounds, named colneleic acid (from linoleic acid) and colnelenic acid (from linolenic acid), could be also produced in potato leaves and tomato roots by rearrangement of 9-hydroperoxides.
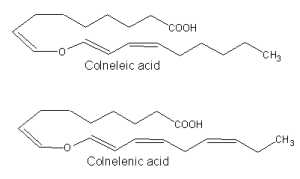
Isomers of colneleic and colnelenic acids were isolated from homogenates of leaves of Clematis vitalba (Ranunculaceae) (Hamberg M, Lipids 2004, 39, 565).
Similarly, 13-lipoxygenase-generated hydroperoxides serve as precursor of other divinyl ether fatty acids which are produced in bulbs of garlic (Grechkin AN et al., FEBS Lett 1995, 371, 159) or Ranunculus leaves (Hamberg M, Lipids 1998, 33, 1061). These compounds were named etheroleic and etherolenic acids.
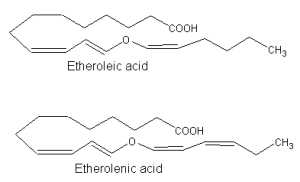
The physiological significance of divinyl ethers is still not fully studied. As infection of potato leaves leads to increased levels of divinyl ether synthase, it was suggested that this pathway could be of importance in the defense of plants against attacking pathogens (Göbel C et al., Biochim Biophys Acta 2002, 1584, 55).
Similar structures have been discovered in the brown alga Laminaria sinclairii, with 18 or 20 carbons and 4, 5 or 6 double bonds (Proteau PJ et al., Lipids 1993, 28, 783), and in the red alga Polyneura latissima, with 20 carbons and 5 double bonds (Jiang ZD et al., Lipids 1997, 32, 231).
![]()
Devenez membre et participez au développement de la Lipidomique au XXIème siècle.
S'inscrire