
LIPOAMINO ACIDS WITH ETHER LINKAGE
These lipids are glycerolipids and are mainly derived from homoserine, they are characterics of algae. Some forms have an alanine moiety instead of homoserine. As the polar head may be considered derived from betaine (N,N,N-trimethyl glycine), these lipids are commonly named betain lipids.
The homoserine-derived lipids were first identified in a yellow-green algae, Ochromonas danica (Chrysophyceae) where they account for more than 50% of total lipids (Brown AE et al., Biochemistry 1974, 13, 3476). It has been suggested that homoserine-derived lipids, which are formed by an ether linkage between homoserine and a diacylglycerol molecule, are widely distributed and even found in some higher plants (Rozentsvet OA et al., Phytochemistry 2000, 54, 401). Furthermore, they were shown to be a substitute for phospholipids in several plants (Brown AE et al., Biochemistry 1974, 13, 3476). This replacement has been well documented in the plants Arabidopsis and oat, and most pronouncedly in the a-proteobacteria Rhodobacter sphaeroides and Sinorhizobium meliloti, which under phosphate stress become depleted of membrane phospholipids and induce the synthesis of non-phosphorous lipids, such as the betaine lipid diacylglyceryl-N,N,N-trimethylhomoserine (Benning C et al., Biochim Biophys Acta 1995, 317, 103). Similar observations were reported for phytoplankton in the ocean using non-phosphorus lipids in response to phosphorus scarcity (Van Mooy BAS et al., Nature 2009, 458, 69).
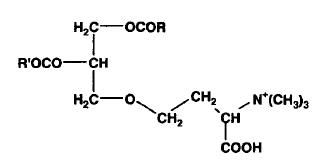
Acyl chains (R1, R2) are 18:0, 18:2(n-6), 18:3(n-6) or (n-3) in Dunaliella (Lynch DV et al., Plant Physiol 1984, 74, 198) and 20:5(n-3), in Chlorella minutissima (Haigh WG et al., Biochim Biophys Acta 1996, 1299, 183) and mainly 18:1(n-9) in Acanthamoeba (Furlong ST et al., J Lipid Res 1986, 27, 1182).
This lipid was shown to be also present in a protozoa, Acanthamoeba, (Furlong ST et al., J Lipid Res 1986, 27, 1182), a fungus (Epidermophyton floccosum) and in several green algae (Volvox, Chlamydomonas, Ulva, Acetabularia, Closterium) but is absent in others (Chlorella, Tetraselmis). It also occurs in mushrooms (Hanus LO et al., J Food Lipids 2008, 15, 370), in vascular cryptogamic plants but not in seed plants (Sato N et al., Phytochemistry 1984, 23, 1625; Kunzler K et al., Phytochemistry 1997, 46, 883).
As they contain a quaternary ammonium group, homoserine-derived lipids resemble phosphatidylcholine in some respects, and a balance between their levels was suggested (Eichenberger W, Plant Sci Lett 1982, 24, 91).
Alanine-derived lipids (diacylglyceryl hydroxymethyltrimethyl-b-alanine) was first identified in Ochromonas danica (Vogel G et al., Chem Phys lipids 1990, 52, 99) and was shown to replace the homoserine-derived lipids in brown algae but are absent in the greens (Eichenberger W, Plant Physiol Biochem 1993, 31, 213).
Another betain lipid, diacylglyceryl carboxyhydroxymethylcholine, was then discovered in Pavlova lutheri (Haptophyceae) (Kato M et al., Phytochemistry 1994, 37, 279).
Lysine-containing diacylglycerol was isolated from Mycobacterium phlei strain IST (Lerouge P et al., Chem Phys Lipids 1988, 49, 161). Lysine is esterified to 1,2- diglyceride via an ester linkage and the major fatty acyl substitutions are palmitic and tuberculostearic acid.
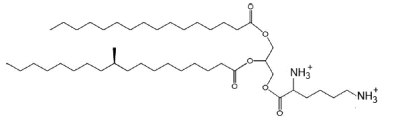
Lysyl diacylglycerol
![]()
Devenez membre et participez au développement de la Lipidomique au XXIème siècle.
S'inscrire