
In his classic studies on fatty acids from pork fat, Chevreul (1823) recognized the nature of oleic acid but it was not prepared in pure condition for a long time. Its structure was not definitively elucidated by a series of complex chemical reactions until later (Baruch J, Ber 1894, 27, 172). Much simpler proof of the now accepted structure of oleic acid was given by means of oxidation techniques (Edmed FG, J Chem Soc 1898, 73, 627). The ozonisation method for the determination of the position of the unsaturated linkage was used for the first time in lipidology in 1903 (Molinari E, Annuario della Soc Chimica di Milano 1903, 9, 507). Oleic acid synthesis was realized in 1934 (Noller CR et al., J Am Chem Soc 1934, 56, 1563).
Mono-unsaturated normal fatty acids are widespread in the living world where they occur mostly as the cis-isomer. Over a hundred naturally occurring monoene fatty acids have been identified. They have the general structure:
CH3(CH2)xCH=CH(CH2)yCOOH
The most frequently they have an even number of carbon atoms and the unique double bond may be in a number of different positions.
The double bond can exist in two stereoisomeric forms :
![]()
The commonest cis-monoenes are of the n-9 series, as oleic acid from olive oil (cis-9-octadecenoic acid) and from quite all seed oils.
Some important monoenoic acids are found below:
|
Systematic name |
Trivial name |
Shorthand designation |
Molecular wt. |
Melting point (°C) |
| cis-4-decenoic | obtusilic | 10:1 (n-6) | 170.3 | |
| cis-9-decenoic | caproleic | 10:1 (n-1) | 170.3 | |
| cis-5-dodecenoic | 5-lauroleic (denticetic) | 12:1 (n-7) | 198.4 | |
| cis-4-dodecenoic | linderic | 12:1 (n-8) | 198.4 | |
| cis-9-tetradecenoic |
myristoleic |
14:1 (n-5) |
226.4 |
– |
| cis-5-tetradecenoic | physeteric | 14:1 (n-9) | 226.4 | |
| cis-4-tetradecenoic | tsuzuic | 14:1 (n-10) | 226.4 | |
| cis-9-hexadecenoic |
palmitoleic |
16:1 (n-7) |
254.4 |
0.5 |
| cis-6-hexadecenoic | sapienic | 16:1 (n-10) | 254.4 | |
| cis-6-octadecenoic |
petroselinic |
18:1 (n-12) |
282.4 |
30 |
| cis-9-octadecenoic |
oleic |
18:1 (n-9) |
282.4 |
16.2 |
| tr-9-octadecenoic | elaidic | tr18:1 (n-9) | 282.4 | 43.7 |
| cis-11-octadecenoic |
vaccenic (asclepic) |
18:1 (n-7) |
282.4 |
39 |
| cis-9-eicosenoic |
gadoleic |
20:1 (n-11) |
310.5 |
25 |
| cis-11-eicosenoic |
gondoic |
20:1 (n-9) |
310.5 |
– |
| cis-11-docosenoic | cetoleic | 22:1 (n-11) | 338.6 | |
| cis-13-docosenoic |
erucic |
22:1 (n-9) |
338.6 |
33.4 |
| cis-15-tetracosenoic |
nervonic |
24:1 (n-9) |
366.6 |
39 |
– Oleic acid is probably the most common fatty acid (olive oil has a high content of this acid: about 60-70%, but it is also found in a large range of nuts). It is found also in seeds such as in Moringa oleifera seeds, a tree cultivated in all tropical and subtropical regions such as Pakistan, Arabia, Central America, North and the South Philippines, Cambodia, Caribbean Islands, and Africa (Saa RW et al., Food Sci Nutr 2019, 7, 1911).
The major sources for human food are palm, rapeseed, soybean, butter, lard and tallow.
While this common fatty acid is mainly found acylated in glycerides, it may be found sometimes as ethyl esters in organs of animal treated with ethanol (Hungund BL et al., J Chem Pharmacol 1988, 37, 3001) and may serve as markers of ethanol intake. (Laposata M, Prog Lipid Res 1998, 37, 307).
The first synthesis of oleic acid appeared in the literature in 1934 (Noller CR et al., J Am Chem Soc 1934, 56, 1563).
Monoethanolamine oleate (ethanolammonium oleate) is a salt formed by the reaction between monoethanolamine and oleic acid. It is used as a sclerosing agent in chronic venous disorders (Rabe E et al., Phlebology. 2014, 29, 338).
Ethyl oleate was identified as a primer pheromone in honey bees in causing a delayed onset of foraging in younger individuals (Leoncini I et al., PNAS 2004, 101, 17559).
In 2010, the Mediterranean diet was recognized by UNESCO as an Intangible Cultural Heritage of Humanity. Olive oil is the most characteristic food of this diet due to its high nutraceutical value and its positive effects have often been attributed to oleic acid (70-80%) but also to its minor component such as oleoylethanolamide with anti-inflammatory and antioxidant effects. That compound derived from oleic acid is an endogenous ligand of the peroxisome proliferator-activated receptor alpha (PPARα) nuclear receptor. Thus, it may regulate dietary fat intake and energy homeostasis (Review in: Santa-María C et al., Nutrients 2023, 15, 224).
– Myristoleic acid is found in most animal depot fats from marine or terrestrial origin. It represents a major component of seed oil from Myristicaceae, some plant species containing up to 30 per cent of this fatty acid in addition to more important amounts of myristic acid. An isomeric form of myristoleic acid was discovered in dolphin and whale head oils (Tsujimoto M, Chem Umschau 1925, 32, 202) and was proved to be the 5-tetradecenoic acid. This uncommon (n-9) fatty acid was more recently described in the retina, acylating an NH2 terminus of a protein (recoverin) related to signal transduction in photoreceptors (Dizhoor AM et al., J Biol Chem 1992, 267, 16033). Furthermore, more recently, the presence of this fatty acid in plasma was shown to be of diagnostic value in patients with defects of long-chain fatty acid oxidation (Onkenhout W et al., Clin Chem 1995, 41, 1467).
– Palmitoleic acid is found mainly in animal fats, particularly in fish and marine mammals. This acid was first noticed in 1854 by Hofstädter PG (Ann 1854, 91, 177) in sperm whale oil and named physetoleic acid. Its exact molecular composition (C16H30O2) was proposed in 1906 by Bull H (Ber 1906, 39, 3570) at the time when its actual name was given by Lewkowitsch. Its structure was established in 1925 (Armstrong EF et al., J Soc Chem Ind 1925, 44, 180T). Marine fish are rich in palmitoleic acid, specifically in the viscera, accounting for almost 11 % of total lipid contents. The manufacture of omega-3 fish oil may also be a good source for extracting that fatty acid (Messina CM et al., Mar Drugs 2021, 19, 160).
It has been determined to be abundant in the Asclepediaceae Asclepis syriaca seed oil (10%) (Chisholm MJ et al., Can J Chem 1960, 38, 2500), in the Australian Macademia ternifolia seed oil (20%) (Bridge RE et al., J Chem Soc 1950, 2396), and in the Connaraceae Roureopsis obliquifoliata (32%) seed oil (Spencer GF et al., JAOCS 1978, 55, 689). Later, it was shown to characterized the seeds of all plants of the Proteaceae family (Badami RC et al., Prog Lipid Res 1981, 19, 119).
The berry oil (not seed oil) of Hippophae rhamnoides (Sea buckthorn) is known to be rich in palmitoleic acid (16-22%) (Dulf F, Chem Cent J 2012, 6, 106). These plants have potential as sustainable feedstocks for producing industrially important octene via metathesis chemistry. 1-Octene is a high-demand feedstock with a global consumption of over half a million tons per year that is primarily used as a comonomer in the expanding production of linear low density polyethylene. Unfortunately, their low yields and poor agronomic properties preclude their commercial use. Metabolic engineering has been done to achieve high levels of n-7 fatty acid accumulation in Arabidopsis (Arabidopsis thaliana) (Nguyen HT et al., Plant Physiol 2010, 154, 1897). The anti-inflammatory effects of palmitoleic acid have been increasingly studied, with research showing it can alleviate macrophage-mediated inflammation induced by palmitic acid or lipopolysaccharide (Souza CO et al., Clin Exp Pharmacol Physiol. 2017; 44, 566). Investigations have shown that palmitoleic acid can exert a protective effect on microglial death induced by palmitic acid via pathways related to pyroptosis and apoptosis (Yu Q et al., Plos One 2024, 19(1), e0297031). Palmitoleic acid was demonstrated to be an adipose tissue-derived hormone (a lipokine) that strongly stimulates in mice muscle insulin action and suppresses hepatosteatosis. Adipose tissue would be able to use lipokines to communicate with distant organs and regulate systemic metabolic homeostasis (Cao H et al., Cell 2008, 134, 933). The amount of palmitoleic acid in serum cholesterol ester reflects the hepatic pool of carbon flux from carbohydrates to fatty acids (Lands W et al., Am J Clin Nutr 1995, 61, 721). Furthermore, as a product of endogenous lipogenesis, palmitoleic acid is reported to correlate with indexes of adiposity and insulin concentrations (Kunesova M et al., Lipids 2002, 37, 27). The present literature seems to suggest that palmitoleic acid is a good marker for excessive carbohydrate consumption, which may reflect a carbohydrate shunt and predict metabolic disease. In fact, an increase in the level of palmitoleic acid around the age of 50 was shown to be associated with an increase in the level of highly sensitive CRP, a marker of inflammation, about twenty years later.
On the other hand, palmitoleic acid has been used as a biomarker to trace dairy products intake in human (Mozaffarian D et al., Am J Clin Nutr 2013, 97, 854).
Sensory evaluation and GC/MS analysis have shown that palmitoleic acid emerged (with geranylacetone and tetradecanoic acid) as one potential compound constituting the quality of female axillary odor for men during the ovulatory phase (Ohgi N et al., iScience 2025, 28, 113087). The fact that hormonal contraception resulted in the loss of these effects suggest that they are dependent of female hormones.
Several researchers have concluded that palmitoleic acid takes part in different mechanisms that improve skin and mucous disorders. Absortion of palmitoleic acid from fish oil was shown to be effective for human skin function in terms of wrinkles, moisture, elasticity, and melanin production and may be useful as a skin nutritional supplement (Sung HK et al., Nutrients 2024, 16(2), 212).
– Petroselinic acid is present as a major component (from 15 to 60%) in many species of Umbelliferae or Apiacae (parsley or coriander oilseeds) and Araliaceae (Hedera helix). Its presence was first noted in 1909 in parsley seed oil (Vongerichten et al., Ber deutsch chem Ges 1909, 42, 1638) and in the seeds of ivy (Hedera) (Steger A et al., Rec Trav Chim 1928, 47, 471) and its exact structure was determined. Within the Umbelliferae, it was shown that the genus Thapsia should be regarded as a useful source of petroselinic acid, which represents an important oleochemical material for the food, cosmetics, chemistry and pharmaceutical industries (Avato P et al., Lipids 2001, 36, 845). Triacylglycerols from T. garganica, T. villosa, T. laciniata, T. minor and T. maxima contain 80–90% of tripetroselinin. Petroselinic acid was also shown to be an important constituent of Geranium sanguineum (Geraniaceae) seed oil (about 48%) (Tsevegsuren N et al., Lipids 2004, 39, 571) and of sweet cicely (Myrrhis odorata, Umbelliferae) (82%). Petroselinic acid was used in the synthesis of new sophorolipids, biosurfactant glycolipid compounds which possess biological activities such as anticancer, antimicrobial, immunoregulatory and antiviral.
The location of the unsaturation at the 6,7-position in petroselinic acid is rare among octadecenoic acids and besides influencing the chemical-physical properties such as its melting point (30 °C), also allows to produce unique derivatives from this molecule.
– Vaccenic acid (from Vacca : cow), named also asclepic acid, was discovered in 1928 (Bertram SH, Biochem Z 1928, 197, 433) in animal fats and butter.
– Gadoleic acid was first noted in cod liver oil (Bull H, Ber 1906, 39, 3570), present in marine fish, it originates from dietary crustacea. Its structure was given in 1933 (Takano M J, Soc Chem Ind Japon 1933, 36, 1317). A shorter fatty acid of the n-11 family, 16:1 n-11, is abundant in psychrophile bacteria, such as Bacillus (Kaneda T, Biochem Biophys Res Commun 1971, 43, 298) and thermophile bacteria, such as Chthonomonas (Vyssotski M et al., Lipids 2011, 46, 1155). cis-9-Docosenoic acid was identified from the dietary seaweed Hizikia fusiformis (hijiki) as a natural sphingomyelin synthesis inhibitor (Sundaraswamy PM et al. , J Agric Food Chem July 2, 2026). These findings demonstrate that gadoleic acid could be a nutraceutical candidate for metabolic disease management.
– Cetoleic acid (22:1 n-11) is present in fish oil, mainly in the species of Gadideae. It must be noticed that cetoleic acid has bioactive properties as it stimulates the synthesis of EPA and DHA from α-linolenic acid in human HepG2 cells and of EPA in salmon hepatocytes in vitro and increases whole-body retention of EPA + DHA in salmon after supplementation in the diet (Østbye TK et al., Br J Nutr 2019, 122, 755). Furthermore, a meta-analysis has suggested that cetoleic acid-rich fish oils could prevent high plasma cholesterol concentration in rodents (Mjaatveit M et al., Br J Nutr 2024, 131, 606). Research on cetoleic acid using cell lines suggests that this fatty acid may positively affect neurological health (review in : Tang PA et al., Lipids Health Dis 16 February 2026).
Pelagia SA has launched a new product (Epax) with cetoleic acid added to marine fish omega-3 fatty acids.
– Nervonic acid, with 24 carbon atoms (24:1n-9), was first believed to be characteristic of fats of Elasmobranch fish (first name: selacholeic acid) where it was discovered (Tsujimoto M, J Soc Chem Ind Japon 1927, 30, 868). In the same year, Klenk E (Z physiol Chem 1927, 166, 287) isolated this fatty acid from cerebrosides of brain tissue, and established its structure. Nervonic acid is particularly abundant in the white matter of animal brains and in peripheral nervous tissue where nervonyl sphingolipids are enriched in the myelin sheath of nerve fibers. Thus, nervonic acid is considered as closely associated with the development and maintenance of the brain and the biosynthesis and improvement of nerve cells (Li Q et al., Food Chem 2019, 301, 125286).
Nervonic acid may promote the repair and regeneration of the damaged nerve cells and tissues (Amminger GP et al., Mol Psychiatry 2012, 17, 1150). Studies have indicated that nervonic acid is useful in the treatment of several neurological diseases, such as Alzheimer’s disease, schizophrenia, psychosis and attention deficit disorder (Wang X et al., Phytomedicine. 2023, 117, 154911), Nervonic acid-enriched diet has been studied and shown to be beneficial for the treatment of genetic disorders of the lipid metabolism, such as Zellweger syndrome or adrenoleukodistrophy. It is also used as a biomarker to predict who will suffer some psychoses (Amminger G P et al., 2012, Nature. 17, 1150). Studies have shown that nervonic acid supplementation is not only beneficial to human health but also can improve the many medical conditions such as neurological diseases, cancers, diabetes, obesity, and their complications (review in: Phung NV et al., Crit Rev Food Sci Nutr 2024, 64, 8766).A review has elaborated on the three primary sources of nervonic acid, discussed its biological functions in neuroprotection, anti-inflammatory activity, and metabolic regulation, and explores its potential applications in biomarker development and functional foods (Li Y et al., Chem Phys Lipids 2025, 273, 105554).
This acid is also present in a Cruciferae (Lunaria annua) in high amounts (22-25%) where it can be suitable (together with erucic acid) for production of high-temperature lubricants and engineering nylons. It is also present (5.2–6.2%) in the seeds of Acer truncatum which are an excellent source of edible oil in China (Wang C et al., Ind Crops Products 2024, 211, 118285), of Tropaeolum speciosum (flame flower), Borago officinalis (borage), and Cannabis sativa (hemp).
Nervonic acid is a prominent fatty acid in the brush border membranes of cod intestine (at ∼50% of the total mono-unsaturated FA) (Gylfason GA et al., Lipid Insights 2012, 5, 19). It was hypothesize that the high cholesterol content and the presence of this rare fatty acid may serve to maintain membrane fluidity in the cold environment.
New strategies of metabolic engineering modifying oleaginous yeast to produce nervonic acid have been described, providing the prospecting views for its production by microbial cells (Liu F et al., J Agric Food Chem 2022, 70, 12772)..
– Erucic acid, with 22 carbon atoms, is found in high amounts (up to 50%) in seed oils of the Cruciferae such as Nasturtium, rape, mustard, Lunaria (38-48%) and of the Tropaeolaceae. Other species have been proposed as sources of erucic acid for nonfood applications, crambe (Crambe abyssinica)(about 60%) and meadowfoam (Limnanthes alba) (about 15%) but there has been no sustained production of these plants. This acid is used in the manufacture of polyethylene film. Tests on animals have shown that ingesting oils containing erucic acid over time can lead to a heart condition called myocardial lipidosis. This is temporary and reversible. Other potential effects observed in animals – including changes in the weight of the liver, kidney and skeletal muscle – occur at slightly higher doses. EFSA’s Panel on Contaminants in the Food Chain established a tolerable daily intake of 7 milligrams per kilogram of body weight (mg/kg bw) per day. For most consumers, especially for toddlers (1-2 years old) and other children (3-10), the main contributors to erucic acid exposure in the diet are pastries, cakes and biscuits. For infants (0-12 months), infant formulae is the main source. Erucic acid is not a safety concern for most consumers as average exposure is less than half the safe level. But it may be a long-term health risk for children up to 10 years of age who consume high amounts of foods containing this substance.
Erucic acid treatment was shown to reduced significantly the tumor burden, glucose levels, and inflammatory and apoptotic markers, and restore the antioxidant levels in 7,12-dimethylbenz[a]anthracene (DMBA)-induced breast cancer in rats (Alamri A et al., Food Bioscience 29 August 2025, 107508). These findings highlight the potential of erucic acid to prevent breast cancer and develop adjunct therapies. Research on erucic acid using cell lines suggests that this fatty acid may positively affect neurological health (review in : Tang PA et al., Lipids Health Dis 16 February 2026)
Brassica oils with a moderate/high content of erucic acid are in fact excellent lubrificants and represent a competitive renewable source for biofuel and biodiesel production. Some improved crops from different brassicas either containing a low content of erucic acid (< 0.5% of total fatty acids) in the seed oil (the so called canola oil, “Canadian oil low in acid”) as well as with very high amounts (nearly 50%) of erucic acid have been developed and are commercially available for industrial applications. The industrial use of erucic acid has considerably increased during the last ten years. The main application is in the production of erucamide, and as starting material to produce behenic acid (C22:0) and caprenin.
– The acid 23:1n-9 occurs almost only in the Holothuroidea (1.6–18% of total FAs), and, therefore, can be considered an important chemotaxonomic marker of this class. Numerous findings indicate that 23:1n-9 is not derived from their diet and is synthesized in their organisms. Kaneniwa has suggested two alternative origins of this fatty acid in animals (Kaneniwa M et al., Comp Biochem Physiol B 1986, 84, 451). First, 23:1n-9 is most likely formed from 24:1n-9 by α-oxidation rather than by desaturation of odd-chain saturated fatty acid. In turn, 24:1 is synthesized from n-9 monoene precursors, including 18:1, 20:1, and 22:1. Monoene n-9 such as 16:1n-9, 18:1n-9, 20:1n-9, and 22:1n-9 are common components of lipids of holothurians and many other marine animals. Therefore, the formation of the 23:1n-9 acid may most likely occur through the elongation of precursors by the following biosynthetic pathway with subsequent α-oxidation: Δ9-18:1 → Δ11-20:1 → Δ13-22:1 → Δ15-24:1→ (α-oxidation) 23:1n-9.
N-1 monoenes : 8-Nonenoic Acid was shown to be emitted by Kocuria flava, a bacterial endophyte of the marine macroalga Bryopsis plumosa. This fatty acid has the property to inhibit the aquaculture pathogen Saprolegnia parasitica (Deutsch Y et al., Mar Drugs 2023, 21, 476). This compound could enable managing oomycete agricultural pathogens in general, and S. parasitica in particular, a major causal agent in aquaculture diseases.
N-3 monoenes : An unusual isoform, the lauroleic acid (9-dodecenoic acid, 12:1 n-3) has been described as a natural metabolite of lauric acid (12:0) in rat hepatocytes (Legrand P et al., Lipids 2002, 37, 569)
N-5 monoenes : Seeds of Androsace septentrionalis (Primulaceae) were shown to contain an unusual fatty acid : 16:1 n-5 (Tsevegsuren N et al., Lipids 2003, 38, 1173). That fatty acid, together with 18:1 n-5, were describeed in deep-sea Foraminifera (Kharlamenko VI et al., Lipids 2017, 52, 345).
N-7 monoenes : The 5-dodecenoic acid (denticetic acid, 12:1 n-7) is present in sperm whale oil. This compound was formerly known to be present in milk lipids.
The rare 21:4 n-7 has been described in some animal species (crab, sponge, protist), but it was found in high amount (about 9 % of the total fatty acids) in a opisthobranch mollusc, Scaphander lignarius (Vasskog T et al., Mar Drugs 2012, 10, 2676). That fatty acid was shown to be cytotoxic against human cancer cell lines normal lung fibroblasts, more than EPA and arachidonic acid.
N-8 monoenes : An isomer (12:1 n-8, linderic acid) has been found as the major fatty acid (47%) in the seed oil of a Lauraceae, Lindera umbellata (Hopkins CY et al., Lipids 1961, 1, 118). This fatty acid is not known to occur in any other plant family.
N-10 monoenes : Sapienic acid (16:1 n-10), a 16-carbon fatty acid with a single cis double bond at the sixth carbon from the carboxyl end, is the most abundant fatty acid in human wax sebum, and among hair-bearing animals is restricted to humans (Nicolaides N, Science 1974, 186, 19). Sapienic acid has been also identified in in human hair and nail samples beside 18:1n-10 (Destaillats F et al., J Chromatogr A 2011, 1218, 9384). Thus this fatty acid is truly unique to sebum, hair and nail and is not found anywhere else in the human body.
Notably, this fatty acid has been implicated in the pathogenesis of acne (Downing DT et al., J Am Acad Dermatol 1986, 14, 221) but was shown to be effective against gram-positive bacteria in human skin sebum (Wille JJ et al.., Skin Pharmacol Physiol 2003, 16, 176). Further works characterized its biosynthesis by a Δ-6 desaturase acting on palmitic acid (Ge L et al., J Invest Dermatol 2003, 120, 707). While sapienic acid is unique to humans in the animal kingdom, it is not unique in life. It is the major fatty acid of certain plant seeds, such as Thunbergia alata of the Acanthaceae family (Spencer GF et al., Lipids 1971, 10, 712). Sapienic acid has been also detected in deep-sea Foraminifera (Kharlamenko VI et al., Lipids 2017, 52, 345).
N-12 fatty acids : 16:1 n-12 has been detected in lipid extracts from deep-sea Foraminifera (Kharlamenko VI et al., Lipids 2017, 52, 345).
N-13 fatty acids : The acid 20:1n-13 is prominent in lipids of the Ophiuroidea (5–24%) and Asteroidea (15–20%), and it is not found in the Crinoidea. The distribution of this lipid depends on the taxonomic position of the species. A high level of 20:1n-13 has been recorded from the coastal brittle star Amphiura elandiformis (8.7%) (Mansour MP et al., Biochem Syst Ecol 2005, 33, 659).
It has been suggested that by GC chromatography the 20:1n-9, reported in other papers on brittle stars and other echinoderms, may be mainly the 20:1n-13 isomer.
N-14 fatty acids : 21:1 n-14 has been detected in lipid extracts from deep-sea Foraminifera (Kharlamenko VI et al., Lipids 2017, 52, 345).
N-15 fatty acids : An unusual 20 carbon fatty acid (20:1 n-15) is found in high concentration (about 60%) in seeds of Limnanthes alba (meadowfoam), an herbaceous winter annual plant native to the pacific Northwest area of the United States.
No fatty acid exists naturally with the double bond close to the terminal methyl group, only one is chemically synthesized from ricinoleic acid, the undecylenic acid (11:1 n-10).
![]()
One geometrical and several positional isomers of oleic acid exist with a trans double bond. Among the naturally occurring trans isomers, the double bond is in the (n-13), (n-12), (n-9) or (n-7) position.
Vegetable oils and fats are almost trans free, if not warmed at high temperature in the presence of active components. In contrast, trans fatty acids occur in most animal fats especially in butter and ruminant fats.
Among the most common, Elaidic acid (t9-octadecenoic acid) and t-Vaccenic acid (t11-octadecenoic acid) are found in the rumen and in lipids of ruminant animals. trans-Vaccenic acid which is the major trans-monounsaturated fatty acid present in several food products (milk, yoghurt, cheese, butter and meats) results from the bio-hydrogenation of rumenic acid.
The elaidinization reaction was first obtained by a French pharmacist and chemist, Jean-Joseph-Étienne Poutet (1779-1858) (Ann Chim Phys 1819, 12, 58), who observed that trioleine could be converted to the consistency of pork lard when treated with the oxides of nitrogen derived from mercurous nitrate (mainly nitrous acid).

Later, Boudet F (Ann Chim Phys 1832, 50, 391; J Pharm 1832, 18, 469) studied accurately the reaction and isolated after saponification of “elaidine” (obtained from triolein) a fatty acid melting at 36°C which he named “acide élaidique” (elaïdic acid), from the Greek name of olive (elais, elaidos).

Until 1952, elaidic acid was known only as a laboratory isomerization product of oleic acid. This trans fatty acid was demonstrated by infrared analysis to be present in substantial quantities in beef fat (Swern D et al., JAOCS 1952, 29, 44). Later, it was shown that trans fatty acids arise in the first stomach of ruminants as products of catalytic hydrogenation of dietary unsaturated fatty acids (conjugated by an isomerase) during bacterial fermentation. As a result, butter, cheese, milk, beef and mutton fats contain approximately 2-8% transfatty acids by weight.
Trans-fatty acids are also formed in varying amounts during the industrial hydrogenation of plant or fish oils. This hydrogenation improves the thermal stability and prevents any oxidative process in very unsaturated oils. The natural kick found in cis-fatty acids disappears and the molecule becomes linear and thus has physical properties similar to saturated fatty acids as it was observed by Poutet in 1819.
Trans-fatty acids are also formed under the action of thiyl radicals.
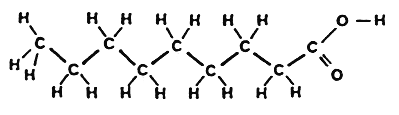
Saturated fatty acids
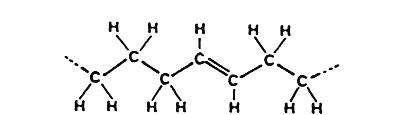
trans-Fatty acid
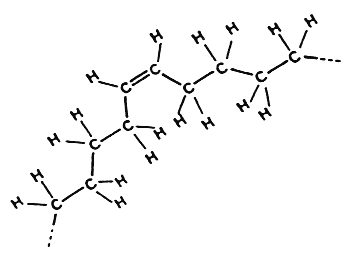
cis-Fatty acid
An unusual trans fatty acid, t3-hexadecenoic acid (trans-16:1 n-13), occurs in eukaryotic photosynthetic membranes (mainly in phosphatidylglycerol) from higher plants and green algae. As this fatty acid is absent from etiolated tissue, it has been inferred that it has a specific role associated with the light reactions of photosynthesis (Gounaris K et al., Biochem J 1986, 237, 313).
trans Fatty acids are formed by some bacteria (predominantly gram negative and under anaerobic conditions) via double-bond migration and isomerization.
The predominant 18:1 trans isomers in partially hydrogenated vegetal oils have their double bond in position t9, t10, t11 and t12, but their distribution (10-22% for t8, t9, t10, t11, t12 or t13 isomer) is distinct from that of milk fat, which contains (2-6% of the total fatty acids) vaccenic acid (t11-18:1) as the predominant isomer (about 60% of t11 and 4-8% for each of the others). The trans-18:1 acid contents of beef meat fat and tallow are about 2 % and 5 %, respectively. Its presence in ruminant fats is explained by a biohydrogenation of linoleic acid occurring in the rumen.
The trans isomers account for about 4.5 % of total fatty acids in ewe milk fat and 3 % in goat milk fat. While the contribution of these two milk sources may be estimated as negligible in most EEC countries, in Greece ewe and goat milk fat contribute for about 45 % of the daily consumption of vaccenic acid.
The daily per capita intake of trans-18:1 acids from ruminant fats was estimated to be about 1.5 g for people from most countries of the EEC, Spain and Portugal being exceptions (about 0.8 g/person/day) (review in Wolff RL, AOCS 1995, 72, 259). An estimation of the trans fatty acid content of foods and intake levels in France has been reported in 2007 (Laloux L et al., Eur J Lipid Sci Technol 2007, 109, 918). The trans fatty acid intakes and their food sources have been determined in the U.S. population (Kris-Etherton PM et al., Lipids 2012, 47, 931).
It is not yet known what effect continuous consumption and exposure to trans fatty acids isomers will have on human health, specifically those present in ruminant fats, deodorized vegetable oils, frying oils, and those present in synthesized products (Aldai N et al., Eur J Lipid Sci technol 2013, 115, 1378). In rats, it was demonstrated that there is a direct retroconversion of dietary trans-vaccenic acid into trans-palmitoleic acid. As it was shown that circulating trans-palmitoleic acid level is associated in man with a lower risk of metabolic syndrome (Kalergis M et al., Front Endocrinol 2013, 4:90), there is a need of deeper knowledge about the origin of that fatty acid and its impact on health. An overview was provided about trans fatty acids origins, their use and consumption among French population with a discussion on their potential human health implications as well as the preventive and regulatory measures undertaken in France (Menas F et al., Eur J Nutr 2013, 52,1289).
The relationships between eighteen‑carbon trans fatty acids and inflammation in the context of atherosclerosis has been reviewed (Valenzuela CA et al., Prog Lipid Res 2019, 76, 101009). Briefly, the evidence associating industrial trans fatty acids with cardiovascular diseases risk factors is fairly consistent, in humans there is a clear a relation between these trans fatty acids and higher levels of inflammatory markers. In contrast, studies in humans, animals and in vitro suggest that ruminant (natural) trans fatty acids have null or mildly beneficial effects in cardiovascular health, metabolic parameters and inflammatory markers.
A review focuses on the mechanisms that underlie the deleterious effects of trans fatty acids by comparing the effects of trans fatty acids to those of cis-unsaturated fatty acids and saturated fatty acids (Oteng AB et al., Adv Nutr 2019 Nov 29). This review also carefully explores the argument that ruminant trans fatty acids have differential effects from industrial trans fatty acids. New data on the molecular mechanisms of action of trans fatty acids may lead to new therapeutic ways for the treatment of diseases characterized by disrupted lipid metabolism. A review has focused on the mechanisms that underlie the deleterious effects of trans fatty acids by juxtaposing effects of trans fatty acids against those of cis-unsaturated fatty acids and saturated fatty acids (Oteng AB et al., Adv Nutr 2020, 11, 697). This review also carefully explores the argument that ruminant trans fatty acids have differential effects from industrial trans fatty acids.
Vaccenic acid has been shown to attenuate complications observed in the metabolic syndrome, including dyslipidemia, fatty liver disease, and low-grade inflammation. An explanation of that property could be that vaccenic acid lowers intestinal inflammation by increasing the production of anandamide and related N-acylethanolamines (Jacome-Sosa M et al., J Lipid Res 2016, 57, 638).
A review of the diversity of adverse health effects of individual trans fatty acid isomers may be consulted (Gebauer SK et al., Lipids 2007, 42, 787). A review of the possible effects of trans fatty acids on heart health and the recommendations for the UK population has been reported (Denny AR, Nutr Bull 2008, 33, 124). While the effects of most of trans fatty acid from ruminants are poorly established, there is increasing evidence that high content of industrial trans fatty acids may cause deleterious effects on human health and life span. A background review paper of their use, consumption, health implications and regulation in France has been published (Menaa F et al., Eur J Nutr 2013, 52, 1289).
DISPERSIVE LIQUID-LIQUID MICROEXTRACTION
Lire la suiteDevenez membre et participez au développement de la Lipidomique au XXIème siècle.
S'inscrire