
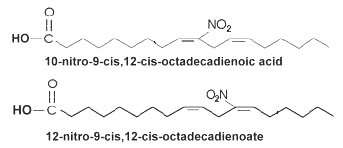
Nitration of unsaturated fatty acids by nitric oxide-derived species (peroxinitrite, nitrous acid …) has been first demonstrated in 1994 for linolenic acid (Rubbo H et al., J Biol Chem 1994, 260, 26066) and then in 1999 for linoleic acid (O’Donnell VB et al., Chem Res Toxicol 1999, 12, 83). It has been shown later that two positional isomers of nitrolinoleic acid (10- and 12-nitro-9,12-octadecadienoic acid) are present in plasma lipoproteins and red cell membranes (about 500 nM, partly esterified to phospholipids or neutral lipids) (Baker PRS et al., PNAS 2004, 101, 11577).
Mass spectrometric analysis of human plasma and urine revealed abundant nitrated derivatives of all principal unsaturated fatty acids. Nitrated palmitoleic, oleic, linoleic, linolenic, arachidonic, eicosapentaenoic and docosahexaenoic acids were detected in concert with their nitrohydroxy derivatives (Baker PR et al., J Biol Chem 2005, 280, 42464). The nitration of linoleic acid is likely produced in vivo by the initiation of its auto-oxidation by nitric oxide (.NO2) by means of hydrogen abstraction from bis-allylic carbon followed by a reaction with .NO2. An acid-catalyzed nitration is also possible during precise physiological or pathological situations or in vitro in the presence of HNO2– radical. mechanisms. Moreover, nitrohydroxyarachidonate has been shown to exhibit vaso-relaxatory effects in vitro, an observation consistent with its production (Balazy M et al., J Pharm Exp Ther 2001, 299, 1). It was shown that nitrated fatty acids are present in plants, specifically in olives and extra virgin olive oil and that an additional formation of NO2-FA from olive oil takes place under acidic gastric conditions (Rubbo H et al., in M. N. Khan et al. (eds.), Nitric Oxide in Plants: Metabolism and Role in Stress Physiology, 2014). Thus, the Mediterranean diet rich in NO2– and unsaturated fatty acids (e.g., linoleic, oleic, and conjugated linoleic), may favor the generation of NO2-fatty acids contributing to the cardiovascular benefits associated with that diet.
It was previously demonstrated that these nitrated fatty acids may serve as cell signaling molecules transducing the vascular actions of .NO (Lim DG et al., PNAS 2002, 99, 15941). Further experiments proved that nitroalkene derivatives of linoleic acid are pluripotent signaling mediators that act directly via receptors and also by transducting the signaling actions of .NO (Schopfer FJ et al., J Biol Chem 2005, 280, 19289). Furthermore, nitrolinoleic acid was shown to be a potent endogenous ligand for peroxisome proliferator-activated receptor gamma (PPARg) (Schopfer FJ et al., PNAS 2005, 102, 2340). Several experimental results provide mechanisms to explain nitric oxide production from nitrated lipids, mechanisms which support the role of lipid sources of nitric oxide in endothelium-independent vasorelaxation (Lima ES et al., Free Rad Biol Med 2005, 39, 532). Nitro-oleic acid was shown to lower blood pressure in a hypertension model in wild-type mice, but was ineffective in mutant mice (Charles RL et al., PNAS 2014, May 19). That protection from hypertension afforded by the oleic acid-rich Mediterranean diet is likely mediated by a nitro-fatty acid-dependent inhibition of soluble epoxide hydrolase, thus preventing the hydrolysis of epoxyeicosatrienoic acid which induces vasodilation.
Several observations have indicated that nitro-linoleic and nitro-oleic acids belong to a class of endogenous anti-inflammatory mediators (Cui T et al., J Biol Chem 2006, 281, 35686). They inhibit cytokine and inducible nitric oxide synthase expression in lipopolysaccharide (LPS) and interferon-g-stimulated monocytes. A nitro-fatty acid (nitro-oleic acid), naturally produced during myocardial ischaemia, was shown to have anti-inflammatory properties and could have therapeutic actions against myocardial ischaemia/reperfusion injury (Rudolph V et al., Cardiovasc Res 2009, cvp275). Later investigations have demonstrated that the anti-inflammatory of nitro-oleic acid may be a consequence of its ability to inhibit irreversibly xanthine oxidoreductase which is able to generate very potent proinflammatory oxidants (Kelley EE et al., J Biol Chem 2008, 283, 36176). Synthesis, properties, and role in biological system of nitrated fatty acids have been reviewed (Rubbo H et al., in Nitric Oxide in Plants: Metabolism and Role in Stress Physiology, M. N. Khan et al. (eds.), Springer International 2014).
Nitro-oleic acid was shown to reduce in vitro the growth and viability of tripple-negative breast cancer cell xenografts in the mammary fat pads of female nude mice (Woodcock CC et al., J Biol Chem. 2017, Nov 20. pii: jbc.M117.814368).
Experiments have demonstrated that NO2-FA are primarily absorbed and transported through the lymphatic system as esterified triacylglycerol species, undergoing initial metabolism in enterocytes regardless of sex, with an unexpected impact on intestinal fatty acid uptake. Thus, assessment of intestinal triacylglycerol uptake by 3H-triolein tracing in mice showed that 10-NO2-oleic acid significantly reduced dietary fat absorption by 75% (Schopfer FJ et al., J Lipid Res 2025, 66, 100855).
Nitrated phospholipids have been recently identified in biological systems and showed to display anti-oxidant and anti-inflammatory potential in models of inflammation in vitro. Using nitrated 1-palmitoyl-2-oleyl-phosphatidyl choline, it has been observed that it was able to induce cellular changes consisting in cytoskeletal rearrangement and cell shrinking, and ultimately, loss of cell adhesion or impaired cell attachment (Duarte S et al., Free Rad Biol Med 2019, 144, 192). Interestingly, the effects of that nitrated phospholipid were not superimposable with those of NO donors, which points to distinctive mechanisms of action but they have shown a stronger parallelism, although not complete overlap, with the effects of nitrated fatty acids. The authors concluded that nitrated phospholipids could be potential novel electrophilic lipid mediators with selective actions.
Reviews on the mechanisms of formation, chemical characterization, and biological properties of nitrated fatty acids may be consulted with interest (Trostchansky A et al., Free Rad Biol Med 2008, 44, 1887; Freeman BA et al., J Biol Chem 2008, 283, 15515; Rubbo H et al., Biochim Biophys Acta 2008, 1780, 1318; Rodriguez L et al., Food Biosci 2026, 75, 108175). The biochemical interactions between nitric oxide and lipid oxidation pathways have reviewed (O’Donnell VB et al., Circ Res 2001, 88, 12). A critical, in-depth analysis of the cardioprotective pathways mediated by nitrite/NO delivered by the Mediterranean diet may be consulted (Nadtochiy SM et al., Nutrition 2011;27(7-8):733). In a review, different synthetic methods and extraction techniques for nitrated fatty acids were described (Elshoura Y et al., Anal Biochem 2024, 694, 115624). Additionally, the reported analytical methods for their characterization and quantification and the associations between their endogenous levels and variable diseases are summarized.
In plants, a transcriptomic analysis by RNA-seq technology established in Arabidopsis a clear signaling role for nitrolinolenic acid, demonstrating that it was involved in plant defense response against different abiotic-stress conditions, mainly by inducing heat shock proteins and supporting a conserved mechanism of action in both animal and plant defense processes. A review of these topics may be consulted (Mata-Pérez C et al., Plant Physiol 2016, 170, 686).
DISPERSIVE LIQUID-LIQUID MICROEXTRACTION
Lire la suiteDevenez membre et participez au développement de la Lipidomique au XXIème siècle.
S'inscrire